Our work focuses on two main questions:
1) How is reproductive diversity maintained in space and time?
2) How do reproductive strategies impact species formation and persistence?
The choice of a mate is vitally important for humans and plants alike. Should you mate at all? If so, how? Where? And who with? These choices have profound impacts for genetics and ecology of an organism's descendants, which ultimately shapes population-level processes and patterns of biodiversity.
We study reproductive diversity in flowering plants because plants have an incredible array of reproductive strategies--asexuality, self-fertilization, and outcrossing (whether within a population, across populations, or hybridization between species). These reproductive strategies can vary within a species, a population, or even among ovules in an individual.
1) How is reproductive diversity maintained in space and time?
2) How do reproductive strategies impact species formation and persistence?
The choice of a mate is vitally important for humans and plants alike. Should you mate at all? If so, how? Where? And who with? These choices have profound impacts for genetics and ecology of an organism's descendants, which ultimately shapes population-level processes and patterns of biodiversity.
We study reproductive diversity in flowering plants because plants have an incredible array of reproductive strategies--asexuality, self-fertilization, and outcrossing (whether within a population, across populations, or hybridization between species). These reproductive strategies can vary within a species, a population, or even among ovules in an individual.
How is reproductive diversity maintained?

Boechera growing in Lemhi County, Idaho
|
The benefits and drawbacks inherent to each form of reproduction depend on the environment in which mating occurs. This means that the consequences of a mating strategy vary across space and time. What environments favor different reproductive strategies, and what ecological benefits and costs do different reproductive strategies incur? In the rock cress Boechera, sexual lineages often self-fertilize (and are thus highly homozygous), while asexuals result from outcrossing and hybridization (thus having high fixed heterozygosity). In some environments, the benefits of self-fertilization (such as more rapid adaptation to novel habitats) may outweigh the drawbacks (such as fitness loss due to inbreeding depression). Similarly, asexual reproduction is hypothesized to carry short-term benefits and long-term costs. |
The evolution of sex in the natural environment
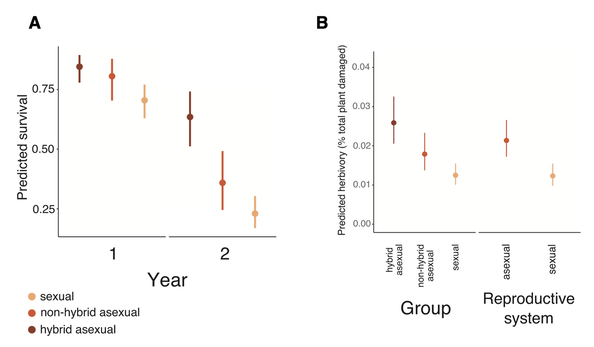
A) Survival of sexual lineages is lower than both hybrid and non-hybrid asexuals. B) Herbivory is elevated in asexuals compared to sexuals, and hybrid asexuality is highest. Both from Rushworth et al. 2020.
Classic evolutionary theory predicts that sexual and asexual reproduction can only coexist in a narrow range of conditions. Yet numerous examples of sex/asex coexistence can be found across plants and animals. Why? In Boechera, ecological differentiation may contribute to largescale sex/asex coexistence (Rushworth et al. 2018).
Hybridization and asexuality have unique effects on fitness in the natural environment. Sexuals have elevated over-winter mortality, likely as a result of inbreeding depression as the most homozygous sexuals have lower fitness. Later in the growing cycle, asexuals--especially hybrid asexuals--experience more insect herbivory than sexuals, which reduces subsequent survival (Rushworth et al. 2020). These results show that both biotic and abiotic factors contribute to the evolution and maintenance of sex in the natural world, and that the costs of sex depend on the identity of your partner and the environment in which sex occurs. Future work will pinpoint specific loci that underlie these complex fitness phenotypes, and the impacts of heterozygosity on their function.
Hybridization and asexuality have unique effects on fitness in the natural environment. Sexuals have elevated over-winter mortality, likely as a result of inbreeding depression as the most homozygous sexuals have lower fitness. Later in the growing cycle, asexuals--especially hybrid asexuals--experience more insect herbivory than sexuals, which reduces subsequent survival (Rushworth et al. 2020). These results show that both biotic and abiotic factors contribute to the evolution and maintenance of sex in the natural world, and that the costs of sex depend on the identity of your partner and the environment in which sex occurs. Future work will pinpoint specific loci that underlie these complex fitness phenotypes, and the impacts of heterozygosity on their function.
Evolution of reproductive isolation in maize and teosinte zones of secondary contact
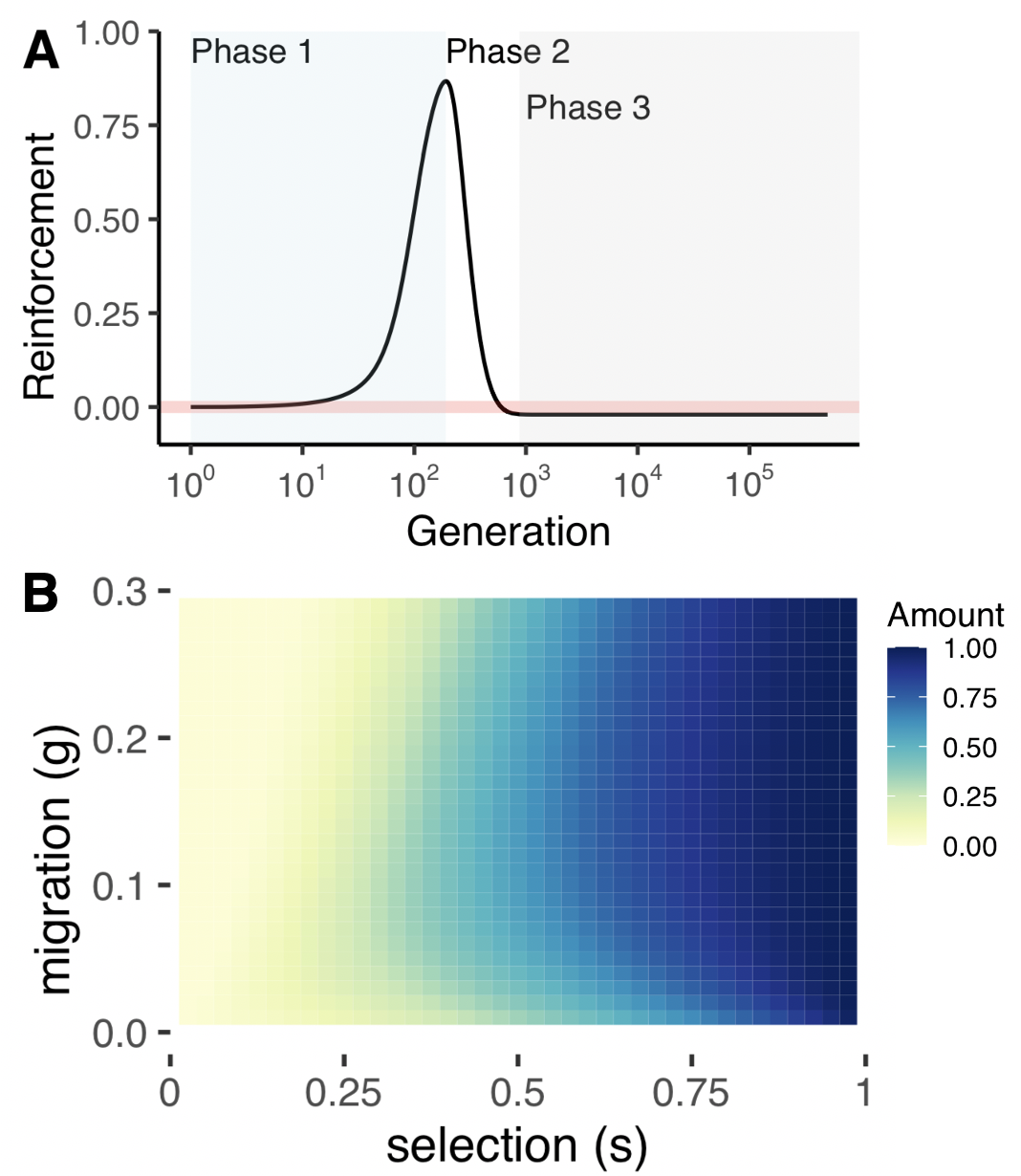
A) Reinforcement dynamics from the model. B) Reinforcement only
reaches completion (dark blue) with exceptionally strong selection.
From Rushworth et al. 2020b.
|
The formation of new species occurs with the evolution of reproductive isolation. In my postdoc with the Brandvain and Ross-Ibarra labs, I used theory to understand the evolutionary forces that result in incomplete reproductive isolation between domesticated maize and its wild progenitor teosinte. Three pairs of loci, called "gametophytic factors," govern postmating prezygotic (PMPZ) isolation via two tightly linked loci: a barrier expressed in silk (stylar) tissue and a pollen-expressed compatibility locus.
In the model we constructed, the mating advantage of the pollen-expressed compatibility allele leads to its spread. Once the compatibility fixes, female choosiness no longer prevents maladaptive matings, and reinforcement subsequently erodes. Reinforcement in this system is almost always transient (Rushworth et al 2022). |
Effects of mating system on reproductive isolation in Boechera
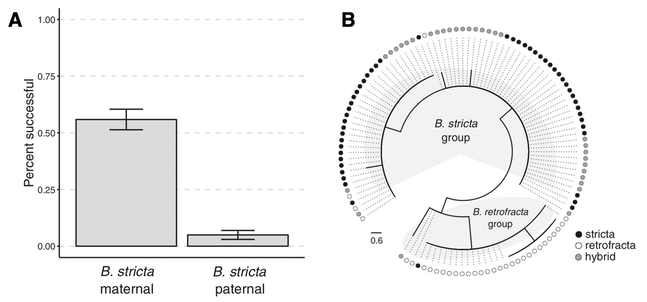
A) PMPZ barriers prevent cross success between B. retrofracta and B. stricta when B. retrofracta acts as the maternal parent. B) A phylogeny of a chloroplast marker trnL shows that wild-collected hybrids also have B. stricta as the maternal parent (hybrids more often cluster with B. stricta, shown in black). From Rushworth and Mitchell-Olds 2020.
How does self-fertilization contribute to species divergence and the evolution of reproductive isolation?
Boechera species hybridize often, but differ in inferred levels of inbreeding. Hybrid inviability, sterility, and cross failure all contribute to reproductive isolation between the two widespread species B. stricta and B. retrofracta. F2 hybrids have lower fitness than their parental lineages, suggesting that, despite their frequent occurrence in the field, not all hybrids are high-fitness asexual lineages. In the greenhouse, crosses fail unless B. stricta is the maternal parent, which is also the pattern in field-collected hybrids (Rushworth and Mitchell-Olds 2020). This result is consistent with the WISO hypothesis (Brandvain and Haig 2005), which we will explore in the future.
How does self-fertilization contribute to species divergence and the evolution of reproductive isolation?
Boechera species hybridize often, but differ in inferred levels of inbreeding. Hybrid inviability, sterility, and cross failure all contribute to reproductive isolation between the two widespread species B. stricta and B. retrofracta. F2 hybrids have lower fitness than their parental lineages, suggesting that, despite their frequent occurrence in the field, not all hybrids are high-fitness asexual lineages. In the greenhouse, crosses fail unless B. stricta is the maternal parent, which is also the pattern in field-collected hybrids (Rushworth and Mitchell-Olds 2020). This result is consistent with the WISO hypothesis (Brandvain and Haig 2005), which we will explore in the future.